viernes, 22 de abril de 2016
MIS BLOGS
Yo tengo tres blogs más que éste:
a través del arte
El blog imaginario
playmundobil
Y mi madre tiene otro blog muy bonito que se llama:
Muñoz Bautista
sábado, 9 de abril de 2016


Aragonito (CaCO3)

El aragonito o aragonita es una de las formas cristalinas del carbonato de calcio (CaCO3), junto con la calcita. Puede encontrarse en forma de estalactitas, y también en la concha de casi todos los moluscos y en el esqueleto de los corales. Entre las variedades del aragonito destaca la llamada flos-ferri (flor de hierro), que se asemeja a un hermoso coral.
El par aragonito/calcita fue el primer caso de polimorfismo mineral reconocido. Esto quiere decir que ambos tienen idéntica composición química, pero diferente estructura cristalina. Debido a esta diferencia, el aragonito es más soluble en agua que la calcita e inestable a temperatura y presión ambientes. De hecho, para periodos geológicos de tiempo (de 10 millones a 100 millones de años), el aragonito tiende a transformarse en calcita. Esta última propiedad puede usarse para determinar la edad de ciertas formaciones rocosas.
El aragonito también pertenece a una serie isomorfa, esto es, un grupo de minerales que pertenecen a la misma clase y presentan la misma estructura cristalina, pero cuya composición es diferente. El aragonito contiene sustituciones isomorfas de bario (witherita), plomo (cerusita), cinc (bromlita) o estroncio (estroncianita). Estos minerales forman el grupo del aragonito.
En cuanto a las aplicaciones del aragonito, son muy limitadas debido a la inestabilidad del mineral. El aragonito sólo suele usarse como piedra ornamental o de coleccionismo.
CRISTALOGRAFÍA
El aragonito cristaliza en el sistema ortorrómbico, en la clase bipiramidal rómbica (2/m 2/m 2/m ó mmm).3 Su grupo espacial es el Pmcn, lo que indica que la celdilla unidad del aragonito es simple o primitiva.
Las dimensiones de la celdilla unidad del aragonito son:
a = 4.96Å, b = 7.97Å, c = 5.74Å; Z = 4.
Agregado de maclas múltiples de aragonito (de unos 4 cm).
En la naturaleza, el aragonito se presenta habitualmente en cristales romboédricos, sencillos o con macla múltiple, que le confieren aspecto de prisma hexagonal (aunque el aragonito no tenga estructura cristalina hexagonal, por eso se llama pseudohexagonal). También formas hialinas coraloides (flos-ferri), fibroso o fibrosoradiado, estalactítico, oolítico y pisolítico.
FORMACIÓN
 El aragonito se forma a partir de aguas termales o géiseres, aguas filtradas que han entrado en contacto con rocas muy calientes situadas a gran profundidad y que han vuelto a emerger a la superficie. Estas aguas disuelven minerales de las rocas a su paso, entre ellos, el calcio. A medida que las aguas termales se evaporan, el calcio que contiene precipita y, cuando entra en contacto con el aire, se combina con el oxígeno y el dióxido de carbono formando los cristales de aragonito.
El aragonito se forma a partir de aguas termales o géiseres, aguas filtradas que han entrado en contacto con rocas muy calientes situadas a gran profundidad y que han vuelto a emerger a la superficie. Estas aguas disuelven minerales de las rocas a su paso, entre ellos, el calcio. A medida que las aguas termales se evaporan, el calcio que contiene precipita y, cuando entra en contacto con el aire, se combina con el oxígeno y el dióxido de carbono formando los cristales de aragonito.
El aragonito puede encontrarse formando estalactitas en cuevas. También puede localizarse en rocas metamórficas o en rocas sedimentarias de los fondos oceánicos, así como en los esqueletos de muchos organismos marinos vivos o recientemente fosilizados. Además, es común en zonas oxidadas de yacimientos metálicos.
Los yacimientos de aragonito más importantes se encuentran en España, entre los que destacan el de la localidad de Luzón (Guadalajara), y los de Minglanilla, en Cuenca, donde se encuentra en una gran variedad de colores. Pueden encontrarse cristales pseudohexagonales en Italia y Sicilia, agregados en piña en Marruecos y variedades estalactíticas y coraloideas en Arizona, Chihuahua (México), Francia (de color azul), Austria e Italia.
En Eslovaquia hay una cueva entera formada de aragonito, llamada la cueva de aragonito Ochtinská. Esta cueva está declarada Patrimonio de la Humanidad por la Unesco, y es una de las tres cuevas de aragonito que se han descubierto en el mundo hasta el momento. Es igualmente destacable en cuanto a sus formaciones excéntricas de aragonito la Cueva de El Soplao en Cantabria (España).
ETIMOLOGÍA E HISTORIA
El nombre aragonito se debe al mineralogista Abraham Gottlob Werner, quien en 1788 lo definió a partir de unos ejemplares procedentes de Molina de Aragón, en Guadalajara, que él atribuyó erróneamente a Aragón.
Habitualmente, los mineros llamaban al aragonito flor de hierro, pues era frecuente encontrarlo en los depósitos de este metal.
Orthoceras

Orthoceras (del griego, orthos, recto y keras, cuerno; "cuerno recto") es un género extinto de moluscos cefalópodos cuya especie tipo, O. regulare, vivió en el Ordovícico hasta el Triásico. Este género se denomina a veces Orthoceratites, y a veces mal denominado como Orthocera u Orthocerus. Probablemente fue pariente del Cameroceras.
| Orthoceras | ||
|---|---|---|
| Rango temporal: 488 Ma-199,6 Ma Ordovícico a Triásico | ||
 Orthoceras regulare | ||
| Taxonomía | ||
| Reino: | Animalia | |
| Filo: | Mollusca | |
| Clase: | Cephalopoda | |
| Subclase: | Nautiloidea | |
| Orden: | Orthocerida† | |
| Familia: | Orthoceratidae† | |
| Género: | Orthoceras† Bruguière, 1789 | |
| Especie: | O. regulare† (Schlotheim, 1820) | |
Trilobites(tres lóbulos)
TRILOBITES


 Tagmas
Tagmas
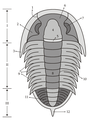 Morfología
Morfología
 Céfalon
Céfalon




TRILOBITES
Los trilobites1 (Trilobita, del latín, "tres lóbulos") son una clase de artrópodos extintos, dentro del subfilo Trilobitomorpha. Son los fósiles más característicos de la Era Paleozoica (Era Primaria). Se han descrito casi 4.000 especies.2
Aparecieron en el período Cámbrico (al inicio del Paleozoico, hace unos 540 millones de años), cuando los organismos ediacaricos ya habían desaparecido, y empezaron a diversificarse ya en el Cámbrico inferior. Tras la extinción masiva de finales del Cámbrico sólo sobrevivieron las formas que habitaban ambientes pelágicos, de aguas profundas. Durante el Ordovícico alcanzaron su máxima diversidad y ocuparon casi todos los nichos ecológicos marinos. A partir del Silúrico presentaron pocos cambios, hasta que en las crisis del Devónico medio y superior sufrieron una importante reducción, extinguiéndose todos los órdenes excepto Proetida. Durante el Carbonífero los representantes del grupo son escasos y restringidos a ambientes de arrecife. Los últimos trilobites, ya sólo habitantes de aguas someras, desaparecieron durante la crisis del límite Permo-Triásico, hace unos 250 millones de años, junto con los euriptéridos o «escorpiones marinos», blastoideos, algunas especies de peces, esponjas, equinodermos, etc. Por tanto, su presencia en la Tierra se prolongó durante todo el Paleozoico, casi 300 millones de años. Los restos fósiles de trilobites son tan abundantes y han sido tan profundamente estudiados, que probablemente sean el grupo de animales extintos más conocido.
Inicialmente se consideraron antepasados de los crustáceos (en especial la cochinilla de humedad terrestre, que comparte ciertas características en común) o incluso de todos los artrópodos (ya que fueron los primeros en aparecer en el registro fósil). Hoy se les considera como un grupo independiente, separados de mandibulados y quelicerados.
| TRILOBITES | ||
|---|---|---|
| Rango temporal: Cámbrico-Pérmico superior | ||
 Fósil de Phacops rana, un trilobites del Devónico | ||
| Taxonomía | ||
| Reino: | Animalia | |
| Filo: | Arthropoda | |
| Subfilo: | Trilobitomorpha† | |
| Clase: | Trilobita† | |
| Órdenes | ||
MORFOLOGÍA
Los trilobites tienen el cuerpo aplanado y liso, más o menos ovalado y dividido en tres Tagmas, céfalon (cefalón), tórax y pigidio; tórax y pigidio forman el tronco. Presentan dos surcos longitudinales que dividen el cuerpo en tres lóbulos claramente delimitados (de donde deriva su nombre): uno central (llamado glabela en el céfalon y raquis en el tronco) y dos laterales (denominados genas o mejillas en el céfalon y pleuras en el tronco). El tegumento dorsal era una gruesa cutícula impregnada de carbonato cálcico, lo que ha facilitado su fosilización. Su tamaño varía desde unos pocos milímetros a más 60 cm en algunas especies gigantes.
MorfologíaCéfalon
CÉFALON
El céfalon ("cabeza") es el resultado de la fusión total de diversos segmentos, y no muestra restos externos de metamerización. Las genas se prolongan por los lados y por detrás por dos puntas genales; las genas están divididas en dos partes por una sutura: una gena fija interna (fixigena) y una gena móvil externa (librigena). La estructura formada por la glabela y las fixigenas se denomina cranidio ("cranidium"). En la cara ventral se localiza el hipostoma, una placa suspendida bajo la glabela que protegía la boca.
Sobre las genas hay un par de grandes ojos compuestos sorprendentemente evolucionados (en algunas especies situados sobre pedúnculos), análogos a los de parientes actuales como los crustáceos e insectos. De hecho, los trilobites fueron los primeros animales en desarrollar ojos complejos, lo que probablemente influyó en su éxito evolutivo. En la parte ventral del céfalon se insertan las antenas unirrámeas, largas y multiarticuladas, y se abre la boca, tras la cual hay tres pares de patas muy similares entre sí. Las antenas son equivalentes, por su posición preoral, a las de los insectos y miriápodos y al primer par de los crustáceos.
TÓRAX
El tórax lo formaban entre 2 y 40 metámeros3 bien diferenciados; están articulados entre sí permitiendo el encabalgamiento de segmentos contiguos con lo que el animal podía enrollarse en forma de bola. Ventralmente, cada metámero lleva un par de patas parecidos entre sí y a las del céfalon.
PIGIDIO
El pigidio forma la parte final del cuerpo y consta de un número variable de metámeros fusionados que forman una estructura compacta. Cada segmento lleva un par de patas similares a las del céfalon y del tórax; a veces, hay un par de cercos apicales multiarticulados.
APÉNDICES
Las patas de los trilobites muestran una sorprendente uniformidad. Usualmente se consideran apéndices birrámeos, que comienzan en una precoxa, de la cual parte un exopodio multiarticulado y con expansiones foliáceas y que probablemente tuviera función branquial y nadadora, y una coxa de la que parte el endopodio, compuesto seis artejos y una uña terminal. Las extremidades estaban protegidas por unas expansiones laterales llamadas lóbulos pleurales, que se extendían hacia afuera desde el lóbulo axial central.
Este tipo de apéndices no son homologables con los apéndices de otros grupos de artrópodos. En los crustáceos que también tienen apéndices birrámeos, la rama externa (exopodio) se inserta en la coxa. Tampoco tienen relación directa con los apéndices de los Merostomados.
PALEOBIOLOGÍA
El desarrollo de los trilobites comportaba una serie de estadios larvales. Las especies más primitivas presentaban un desarrollo larvario completo mientras que en las posteriores el proceso se simplificaba.
El primer estadio larval se conoce como protaspis, formado básicamente por el céfalon y el protopigidio. A éste seguía el estadio meraspis en el que se diferenciaban ya algunos segmentos del tórax y el pigidio. El tercer período u holaspis comprende las larvas que ya han adquirido la metamerización completa, pero son aún mucho más pequeñas que un adulto, al que se llegaba después de una serie de mudas.
PALEOECOLOGÍA
Exclusivamente marinos, estaban totalmente ausentes de ambientes de agua dulce y salobre; por su forma aplanada, ojos en posición dorsal y dureza de la cara dorsal se deduce que la mayoría eran animales bentónicos. Seguramente eran micrófagos, filtrando el barro del fondo en que vivían para obtener el alimento, ya que carecían de apéndices excavadores o prénsiles, así como de piezas bucales trituradoras. Algunas especies se hicieron secundariamente pelágicas y desarrollaron expansiones espinosas para favorecer la flotabilidad. Estas espinas también estaban presentes en las larvas protaspis, que son, por tanto, consideradas formas pelágicas con un gran potencial colonizador.
La reducción y pérdida de los ojos experimentada por diversas especies está relacionada seguramente con una adaptación a la zona afótica y la colonización de aguas profundas.
TAFONOMÍA
Como consecuencia de las mudas es muy frecuente el hallazgo de exuvias desarticuladas en el registro fósil, sobre todo cefalones —o cranidios y mejillas librígenas por separado— y pigidios (un mismo individuo puede producir indicios múltiples de su existencia).
ICNOFÓSLES
A la actividad biológica de los trilobites se atribuyen, principalmente, los icnofósiles Cruziana, Diplichnites y Rusophycus que, por otra parte, son también realizados por otros organismos.4 Normalmente se encuentran como contramoldes, en relieve invertido, en la base (cara inferior) de capas de arenisca o cuarcita.
Cruziana se interpreta como pistas fósiles debidas a la locomoción y la alimentación (repichnia y pascichnia) de trilobites en el fondo arenoso, e incluye, dada la gran diversidad de Trilobita, numerosas paraespecies. Son pistas longitudinales que en epirelieve aparecen formadas por dos surcos que dejan una cresta central, en ocasiones con otros dos surcos más pequeños laterales, y en hiporelieves como dos crestas longitudinales con un surco central. En ambos casos aparecen finas estriaciones oblícuas en forma de "V" —que indican el sentido contrario al del avance— formadas por los apéndices. También han recibido el nombre informal de "Bilobites", actualmente en desuso.5
Diplichnites se interpreta como pistas del desplazamiento rápido (repichnia) sobre la superficie arenosa. Aparece como dos pistas paralelas formadas por multitud de pequeñas huellas estrechamente espaciadas entre sí. Las pequeñas huellas se corresponden con los apéndices del organismo y su profundidad y distribución indican las condiciones de este desplazamiento. En ocasiones las huellas aparecen como finas estriaciones cuando el organismo sufrió un desplazamiento lateral debido a corrientes marinas y a veces evoluciona hacia rastros tipo Cruziana a través de formas de transición.6
Rusophycus se interpreta como huellas de reposo (cubichnia) o quizás de alimentación. Se presentan como dos surcos cortos y finamente estriados, más profundos que Cruziana y por lo general aislados de otras pistas. Son resultado de un proceso de excavación o enterramiento del organismo productor para su ocultamiento o acecho en el que se marcan dos lóbulos estriados formados por el movimiento de los dos grupos de apéndices.
PRIMEROS TRILOBITES
Los trilobites más antiguos que se conocen del registro fósil son fallotaspidos y redlichiidos, ambos del órden Redlichiida, y bigotínidos (orden Ptychoparida, superfamília Ellipsocephaloidea), datados de 540 a 520 millones de años.8 9 Los primeros trilobites incluyen a Profallotaspis jakutensis (Siberia), Fritzaspis spp. (oeste de los Estados Unidos), Hupetina antiqua (Marruecos)10 y Serrania gordaensi (España).11 Se piensa que los trilobites se originaron en lo que hoy es Siberia, con posterior distribución y radiación desde esta región.8
Fósil de Caracol

Hay muchas clases de fósiles. Los más comunes son restos de ammonoidea. Muchos de ellos muestran todos los detalles originales del caracol o del hueso, incluso examinados al microscopio. Los poros y otros espacios pequeños en su estructura se llenan de minerales.
Por ejemplo, un caracol puede disolverse totalmente después de quedar enterrado. La impresión que queda en la roca puede llenarse con otro material y formar una réplica exacta del caracol. En otros casos, el caracol se disuelve y tan sólo queda el hueco en la piedra, una especie de molde que los paleontólogos pueden llenar con yeso para descubrir cómo se veía el animal.
Escalas de minerales: dureza, mohs, rosiwal, knoop
ESCALA DE DUREZA
La dureza es la oposición que ofrecen los materiales a alteraciones como la penetración, la abrasión, el rayado, la cortadura, y las deformaciones permanentes entre otras. En el transcurso de la historia, durante el estudio y clasificación de los minerales, hubo un momento en que se hacía pertinente establecer un método que permitiera discernir los diferentes grados de dureza de las rocas y minerales. El primer intento de establecer un procedimiento para tal fin, poco científico, pero en la práctica bastante profesional, se debió a Friedrich Mohs. Su sencillez (tanto de memorización como de aplicación), lo ha afianzado en esta posición, ya que puede emplearse en el trabajo de campo de los geólogos.
Al tratar de establecer comparaciones de dureza con valores absolutos y más precisos, se crearon otros métodos y escalas que, por lo general, adoptaron el nombre de su creador. El presente artículo recoge dichas escalas.
ESCALA DE MOHS
La dureza es la oposición que ofrecen los materiales a alteraciones como la penetración, la abrasión, el rayado, la cortadura, y las deformaciones permanentes entre otras. En el transcurso de la historia, durante el estudio y clasificación de los minerales, hubo un momento en que se hacía pertinente establecer un método que permitiera discernir los diferentes grados de dureza de las rocas y minerales. El primer intento de establecer un procedimiento para tal fin, poco científico, pero en la práctica bastante profesional, se debió a Friedrich Mohs. Su sencillez (tanto de memorización como de aplicación), lo ha afianzado en esta posición, ya que puede emplearse en el trabajo de campo de los geólogos.
Al tratar de establecer comparaciones de dureza con valores absolutos y más precisos, se crearon otros métodos y escalas que, por lo general, adoptaron el nombre de su creador. El presente artículo recoge dichas escalas.
El 16 de agosto de 2012, investigadores del Carnegie Institute of Technology inventaron un nuevo alótropo del carbono, más duro que el diamante, capaz de mantenerse estable en condiciones normales, aunque se obtenía a partir de un material formado por esferas de carbono-60, sometiéndolo a una presión de 320.000 atm.1

ESCALA DE ROSIWAL
La escala de Rosiwal debe su nombre al ilustre geólogo austriaco August Karl Rosiwal. La escala Rosiwal basa su medición en valores absolutos, a diferencia de la escala de Mohs cuyos valores relativos son más apropiados para la investigación de campo (in situ).
Mide en escala absoluta la dureza de los minerales. Se expresa como la resistencia a la abrasión medida en pruebas de laboratorio tomando como base el corindón con un valor de 1000.
ESCALA DE KNOOP
El test de dureza de Knoop (pronunciado ku-nūp) es una prueba de microdureza, un examen realizado para determinar la dureza mecánica especialmente de materiales muy quebradizos o láminas finas, donde solo se pueden hacer hendiduras pequeñas para realizar la prueba. La prueba fue desarrollada por Frederick Knoop y sus colegas del National Bureau of Standards (actualmente el NIST) de EE. UU. en 1939,2 y fue definido por el estándar de la ASTM D1474.

Fósil de Coral

CORAL
Los corales marinos son animales coloniales, salvo excepciones,nota 1 pertenecientes al filo Cnidaria, clase Anthozoa. Las colonias están formadas por hasta miles de individuos zooides y pueden alcanzar grandes dimensiones.
Aunque los corales pueden atrapar plancton y pequeños peces con las células urticantes en sus tentáculos, la mayoría de los corales obtienen la mayor parte de sus nutrientes de las algas unicelulares fotosintéticas denominadas zooxantela, que viven dentro del tejido del coral. Estos corales requieren de luz solar y crecen en agua clara y poco profunda, normalmente a profundidades menores de 60 metros. Los corales pueden ser los principales contribuyentes a la estructura física de los arrecifes de coral que se formaron en aguas tropicales y subtropicales, como la enorme Gran Barrera de Coral en Australia y el arrecife Mesoamericano en el mar Caribe. Otros corales, que no tienen una relación simbiótica con algas, pueden vivir en aguas mucho más profundas y en temperaturas mucha más bajas, como las especies del género Lophelia que pueden sobrevivir hasta una profundidad de 3000 metros.2
El término "coral" no tiene ningún significado taxonómico y es poco preciso; suele usarse para designar a los antozoos en general, tanto a los que generan un esqueleto calcáreo duro, especialmente los que construyen colonias ramificadas, como las acroporas; pero también es común denominar coral a especies con colonias compactas ("coral cerebro" como Lobophyllia) e incluso con esqueleto córneo y flexible, como las gorgonias. Asimismo, se llaman corales blandos a las especies del orden Alcyonacea, que no generan esqueleto y utilizan el calcio en forma de espículas repartidas por su tejido carnoso, para proporcionarles mayor rigidez y consistencia.
Tanto en el mundo del buceo como en acuariofilia, los corales se dividen en blandos y duros, según tengan esqueleto o no. Y los duros, a su vez, se subdividen en duros de pólipo corto y duros de pólipo largo, small polyp stony (SPS) y large polyp stony (LPS), en inglés. Esta clasificación se refiere al tamaño del pólipo del género, que en los SPS como Montipora, suele ser de 1 o 2 mm3 y en los LPS, como Heliofungia actiniformis de hasta 20 cm.4 Pero esta división es poco científica y genera no pocas excepciones bajo un análisis riguroso de las diversas especies. La comunidad científica se refiere a micropólipos cuando los pólipos coralinos tienen entre 1 y 2 mm de diámetro, y alude a macropólipos para aquellos pólipos entre 10 y 40 mm de diámetro. No obstante, la gran mayoría de los corales de todos los arrecifes del mundo tienen pólipos con un diámetro entre 2 y 10 mm, justo entre esas categorías.
| Coral | ||
|---|---|---|
 Coral en forma de pilar (Dendrogyra cylindricus) | ||
| Taxonomía | ||
| Reino: | Animalia | |
| Filo: | Cnidaria | |
| Clase: | Anthozoa Ehrenberg, 1831 | |
| Subclases y órdenes existentes | ||
Subclase: Octocorallia
Orden: Alcyonacea Orden: Helioporacea Orden: Pennatulacea Subclase: Hexacorallia Orden: Antipatharia Orden: Ceriantharia Orden: Corallimorpharia Orden: Scleractinia Orden: Zoanthidea 1 Véase también Anthozoa | ||
Diente de Tiburón

¿CÓMO PODEMOS SABER SI NUESTROS DIENTES DE TIBURÓN SON REALES?
Los tiburones pierden miles de dientes mientras están vivos, ya que les crecen constantemente otros nuevos. Sus dientes, siendo más resistente que sus esqueletos cartilaginosos, permanecen en el ambiente mucho tiempo después de que se hayan salido o de que el tiburón muera, y algunos se fosilizan. Esto significa que no hay escasez de verdaderos dientes de tiburón antiguos y nuevos en el mundo. Sin embargo, sí existen las falsificaciones. En algunos casos, tales como los dientes nuevos de especies amenazadas como el gran tiburón blanco, es de hecho mejor comprar falsificaciones. En otras situaciones, puedes decepcionarte al descubrir que el "diente de tiburón fosilizado" que compraste en realidad está hecho de plástico. Notar la diferencia es fácil en la mayoría de los casos. Los dientes de tiburón no son tan valiosos como para que muchos vendedores gasten tiempo y dinero en crear falsificaciones de alta calidad. La mayoría de los dientes de tiburón falsos se venden abiertamente como tales, y los reales no suelen valer mucho más.
miércoles, 6 de abril de 2016
Ammonite
EL AMMONITE


AMMONOIDEA
Los ammonoideos (Ammonoidea), conocidos comúnmente como amonites,1 son una subclase de moluscos cefalópodos extintos que existieron en los mares desde el Ordovícico (hace unos 340 millones de años) hasta finales del Cretácico (hace 66 millones de años).2 Gracias a su rápida evolución y distribución mundial son excelentes fósiles guía para la datación de rocas y han posibilitado la elaboración de sucesiones de biozonas de gran precisión bioestratigráfica.
ANATOMÍA
Orientación de la concha:
La región anterior es la que se sitúa hacia la abertura de la concha, la región posterior es la contraria a la abertura. La región ventral se considera la parte externa de las vueltas de espira de la concha, mientras que la región dorsal es la parte interior. La concha de un amonites adulto podía llegar a ser del tamaño de la rueda de un camión.
Partes blandas
Al ser conocidos solo por sus restos fósiles, poco se sabe de las partes blandas del organismo: se supone que eran parecidos a los actuales nautilos, con una corona de tentáculos en la cabeza que asoman por la abertura de la concha. El cuerpo blando se alojaba en la cámara de habitación, la cámara final y de mayor tamaño de la concha.
Partes duras
Poseían una concha fina de aragonito, planiespiralada y con una ornamentación muy marcada, constituida por costillas bien definidas. Dicha concha se divide en dos zonas: el "fragmocono", que es la parte tabicada de la concha donde se almacenan los gases que controlan la flotación del animal; y la "cámara interior", que es donde se alojaban las partes blandas.
En el interior del "fragmocono" se encuentran los "septos", que tabican esta parte de la concha, tienen su concavidad dirigida hacia la región anterior. Los amonites eran pues organismos opistocélicos. Los septos poseen un orificio, llamado "gollete sifonal", orientado hacia la región posterior que permite la comunicación con el resto de septos a través del sifón. La intersección de los septos con la pared externa del fragmocono dibuja en ella la llamada "sutura".
Existen varios tipos de suturas:
Ortoceratítica: simple, con una pequeña inflexión
Goniatítica: con inflexiones más marcadas y redondeadas.
Agoniatítica: las inflexiones son puntiagudas.
Ceratítica: inflexiones con forma de herradura y crenuladas alternativamente.
Amonítica: las inflexiones están muy recortadas y con múltiples subdivisiones.
Sección de un ammonoideo mostrando las cámaras con diferentes rellenos sedimentarios y crecimientos minerales.
La ventaja directa de la complicación de la sutura está relacionada con el aumento de resistencia de la concha. Cuanto más contacto hay entre el tabique y la pared del fragmocono, más resistente será éste.
Esta ventaja no fue aprovechada por los ammonoideos para poder descender a mayores profundidades, sino que vino acompañada por una disminución del grosor de la concha, perdiendo así peso pero no resistencia y ganando efectividad natatoria. El que no aparecieran especies que pudiesen bajar a mayores profundidades está relacionado con la presión interna del sifón, ya que era menor que la del fluido del exterior, por lo que este sifón explotaría si profundizaran.
Para definir la morfología de la concha debemos fijarnos en la sección de la espiral, que puede ser lanceolada, semicircular o circular, y en el grado de abrazamiento de las espiras, que puede ser oxicono, serpenticono o platicono.
Ornamentación
Arietites bucklandi.
Las costillas se encuentran tanto en el fragmocono como en la cámara de habitación, mientras que solo hay suturas en el fragmocono. La ornamentación puede llegar a ser muy compleja. Deberían ser estructuras que facilitasen el camuflaje y diesen resistencia a la concha (costillas anchas) o bien disminuyesen el rozamiento de la concha (costillas finas).
Carena
Es un ligero engrosamiento de la parte ventral. Se trata de una estructura que funcionaría a modo de quilla de barco, lo que mejoraría la natación.
No todos los amonites poseen carena, y cuando la tienen se puede dar el caso de que el sifón se encuentre o no dentro de ella. Si el sifón pasa por la carena, no suele conservarse en el molde interno; pero sí se conserva si el sifón no pasa por ella. Como el sifón no alcanza la cámara de habitación en ninguno de los casos, la carena, si existe, se observara siempre en la cámara de habitación.
Estructuras peristomales
Son estructuras que bordean el orificio de la cámara de habitación. En los nautilos, cuando existen, son muy sencillas; en los ammonoideos en cambio son más complejas.
Posiblemente sean diferenciadores sexuales, siendo exclusivos de los machos. También actuarían como protectores de las partes blandas y ayudarían al alcance del equilibrio hidrostático.
Aptychus
Es una estructura exclusiva de los amonites. Es un tipo de opérculo calcificado que se situaba cerca de la abertura de la concha y funcionaba a modo de “puerta”, protegiendo las partes blandas del individuo una vez cerrada. Está compuesto por un par de piezas simétricas de calcita, que tienen la apariencia de las valvas de un bivalvo, de las que se diferencia porque las estrías de crecimiento se encuentran en la parte cóncava. En algunos amonites los aptychus se encuentran dentro de las cámaras de habitación, en cuyo caso serían estructuras masticadoras de individuos malacófagos (comedores de organismos con concha).
Son difíciles de encontrar al lado de la concha de los amonites al que pertenecieron debido a que la concha seguiría flotando una vez muerto el organismo, mientras que el aptychus se desprendería al descomponerse las partes blandas. Por esta razón se clasificaron como un taxón independiente, porque no se puede saber a qué género de amonites pertenecen.
PALEOECOLOGÍA
Para estudiar el ambiente que habitaban las diferentes especies de amonites hay que estudiar las posiciones del centro de gravedad y de flotación de la concha, ya que éstos determinan la forma de desplazamiento:
La longitud de la cámara de habitación determina la posición del centro de gravedad.
Estabilidad estática: está en relación directa con la distancia entre los centros de flotación y gravedad.
Estabilidad dinámica: depende de la forma de la cámara de habitación; básicamente, de la distancia entre la abertura de la concha (punto de empuje del individuo) y el centro de flotación.
Gracias a estas características, medibles en las conchas fósiles, se puede determinar si se trataba de buenos o malos nadadores, lo que los restringe a un medio marino determinado: los malos nadadores vivirían principalmente en los fondos marinos y cerca de la costa, mientras que los buenos nadadores vivirían en mar abierto.
CLASIFICACIÓN
Principales patrones de suturas de los tabiques en amonites y su distribución en los diferentes órdenes.
La subclase Ammonoidea si divide en cinco órdenes:[cita requerida]
Orden Goniatitida
Orden Ceratitida
Orden Phylloceratida
Orden Lytoceratida
Orden Ammonitida
Superfamilia Hoplitaceae
Superfamilia Perisphinctaceae
Superfamilia Stephanocerataceae
Superfamilia Spirocerataceae
Superfamilia Psilocerataceae
Superfamilia Eoderocerataceae
Superfamilia Hildocerataceae
Entre los más famosos: la Geoda de Amatista
AMATISTA

FORMACIÓN



La amatista es una variedad macrocristalina del cuarzo. Su color violeta característico puede ser más o menos intenso, según la cantidad de hierro (Fe+3) que contenga. Puede presentarse coloreada por zonas con cuarzo transparente o amarillo. Las puntas suelen ser más oscuras o degradarse hasta el cuarzo incoloro.
A pesar de que es muy resistente a los ácidos, la amatista es muy susceptible al calor. De hecho, al calentarla a más de 300 °C cambia su color a café pardo, amarillo, anaranjado o verde, según su calidad y lugar de origen:
450 °C: se vuelve amarilla
500 °C: toma un color anaranjado fuerte (amatista quemada)
600 °C: se vuelve muy lechosa
Estos cambios en la coloración se deben a los cambios en la valencia del hierro que contiene, entre otras cosas. Se puede recuperar el color original de la amatista sometiéndola a irradiaciones. Se diferencia de otras piedras tratadas al calor en que presenta un dicroísmo púrpura azulado y púrpura rojizo.
No tiene una absorción del espectro lumínico característica. Suele presentar inclusiones en forma de marcas paralelas, conocidas como rayas de cebra y arañazos de tigre, causadas por maclas romboédricas.
| Amatista | ||
|---|---|---|
 | ||
| General | ||
| Categoría | Mineral | |
| Clase | Silicatos | |
| Fórmula química | SiO2::Fe+3 | |
| Propiedades físicas | ||
| Color | Violeta en diversas tonalidades | |
| Raya | Blanca | |
| Lustre | Entre céreo y vítreo | |
| Transparencia | Transparente a translúcida | |
| Sistema cristalino | Trigonal | |
| Hábito cristalino | Prisma + romboedro, trapezoedro, bipirámide | |
| Exfoliación | No | |
| Fractura | Concoidea | |
| Dureza | 7 | |
| Peso específico | 2,65 constante; variable en variedades con compuestos agregados | |
| Densidad | 2,65 | |
| Índice de refracción | nω = 1,543 - 1,553 nε = 1,552 - 1,554 | |
| Punto de fusión | 1650 (±75) °C | |
| Solubilidad | Insoluble al H2O | |
| Otras características | La amatista es una variedad del cuarzo | |
| Minerales relacionados | ||
| Cuarzo, Bolivianita o Ametrino, Ayoreita, Ágata | ||
FORMACIÓN
La amatista es un mineral de origen magmático. Se forma en filones con soluciones ricas en óxidos de hierro, que le dan su color morado característico a temperaturas inferiores a los 300 °C. Lo más habitual es encontrar la amatista tapizando el interior de ágatas en forma de geodas, a veces gigantescas. También se pueden encontrar en forma de drusas (cristales que recubren la superficie de una piedra) o en filones, acompañada de otros minerales.
Los principales yacimientos se encuentran en los Urales, Alemania, oeste de Australia, Zambia, Brasil, departamento de Artigas en Uruguay, Estados Unidos, Canadá, India, Sri Lanka, Bolivia, España, Argentina y en Túnez.
JOYERÍA
La amatista es la variedad del cuarzo más apreciada. Las amatistas más perfectas se tallan para joyería, y el resto se utiliza para hacer objetos de arte. Tradicionalmente, la amatista se incluía entre las piedras preciosas más valoradas (junto con el diamante, el rubí, el zafiro y la esmeralda).
Debido a su importancia como gema, existen varios términos utilizados en joyería para describir las distintas tonalidades de la amatista. “Rosa de Francia” se refiere a la amatista de color lila claro, mientras que “Siberiana” es la amatista de color violeta intenso con destellos rojos, la variedad más preciada. También existen gemas que son una mezcla natural entre amatista y citrino, a las que se les ha dado el nombre de “ametrino” o “bolivianita”. Por último, la amatista calentada, que adquiere una tonalidad amarillenta, se suele comercializar como citrino (una variedad de cuarzo de color ámbar muy escasa al natural).
ETIMOLOGÍA E HISTORIA
El nombre ‘amatista’ proviene del griego amethystos (no borracho), ya que esta piedra era considerada un potente antídoto contra la embriaguez. Según la mitología griega, Dioniso, dios del vino y el desenfreno, pretendía a una doncella llamada Amethystos, la cual deseaba permanecer casta. La diosa Artemisa escuchó sus plegarias, y transformó a la mujer en una roca blanca. Dioniso, humillado, vertió vino sobre la roca a modo de disculpa, tiñendo así de púrpura los cristales.
La amatista se conoce desde hace miles de años, pues ya en el antiguo Egipto se utilizaba para crear joyas, sellos personales y tallas. En la Edad Media, el cristianismo adoptó la amatista como símbolo de renuncia a los bienes terrenales y castidad, y aún hoy la llevan en forma de anillos muchos cardenales y obispos. La amatista simboliza además la sabiduría divina.
Suscribirse a:
Entradas (Atom)